








Уважаемая Марианна Ильинична, спасибо огромное за этот пост. Но текст обрывается на полуслове. Подскажите, пожалуйста, где его можно дочитать (если он где-то существует). Какую литературу по нейрофизиологии вы можете порекомендовать?
Морфофункциональное строение нервной системы.
 Нейроны — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов.
Нейроны — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов.
Нейронные связи, сформированные на ранней стадии развития мозга, продолжают изменяться на протяжении всего периода жизни человека. Уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний — синапсов. Нейроны не способны к делению, но генетически запрограммированы на образование связей. Связи образуются благодаря синапсам. Чем больше синапсов, тем больше связей, тем выше показатели ВПФ.
Все нейроны функционально делятся на афферентные (импульсы к центру), эфферентные (импульсы от центра),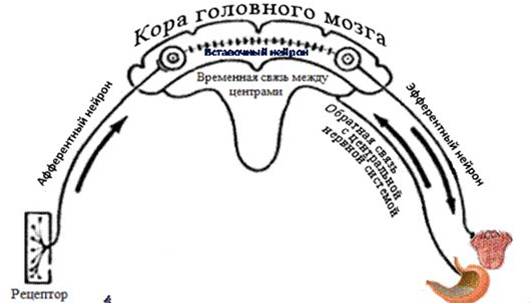 вставочные (предварительная переработка импульсов, образование коллатералей, бывают тормозящими и возбуждающими), рецепторные (от рецепторов в ЦНС, они находятся за пределами ЦНС, в корешках спинномозговых или черепных нервов).
вставочные (предварительная переработка импульсов, образование коллатералей, бывают тормозящими и возбуждающими), рецепторные (от рецепторов в ЦНС, они находятся за пределами ЦНС, в корешках спинномозговых или черепных нервов).
Ядро нейрона содержит генетический материал. Генетический аппарат обеспечивает дифференцировку, конечную форму клетки, а также типичные для данной клетки связи. Другой существенной функцией ядра является регуляция синтеза белка нейрона в течение всей его жизни. Деятельное состояние нейронов сопровождается трофическими процессами—усилением в них синтеза белков. При различных воздействиях, вызывающих возбуждение нервных клеток, в том числе при мышечной тренировке, в их ткани значительно возрастает количество белка и РНК, при тормозных же состояниях и утомлении нейронов содержание этих веществ уменьшается. В процессе восстановления оно возвращается к исходному уровню или превышает его. Часть синтезированного в нейроне белка компенсирует его расходы в теле клетки во время деятельности, а другая часть перемещается вдоль по аксону (со скоростью около 1—3 мм в сутки) и, вероятно, участвует в биохимических процессах в синапсах.
В митохондриях нейронов преобразуются энергия для работы. Резкие изменения митохондрий вплоть до разрушения, а следовательно, и угнетение деятельности нейронов отмечаются при различных неблагоприятных воздействиях (длительном торможении в центральной нервной системе, при интенсивном рентгеновском облучении, кислородном голодании и гипотермии).
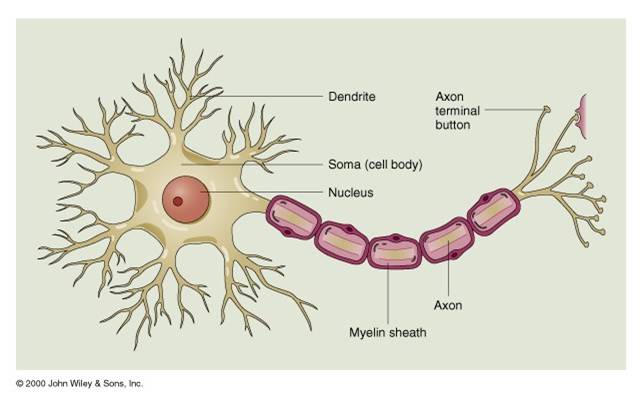
Клеточные скопления образуют серое вещество мозга. Между ядрами, группами клеток и между отдельными клетками проходят миелинизированные или немиелинизированные волокна: аксоны и дендриты.
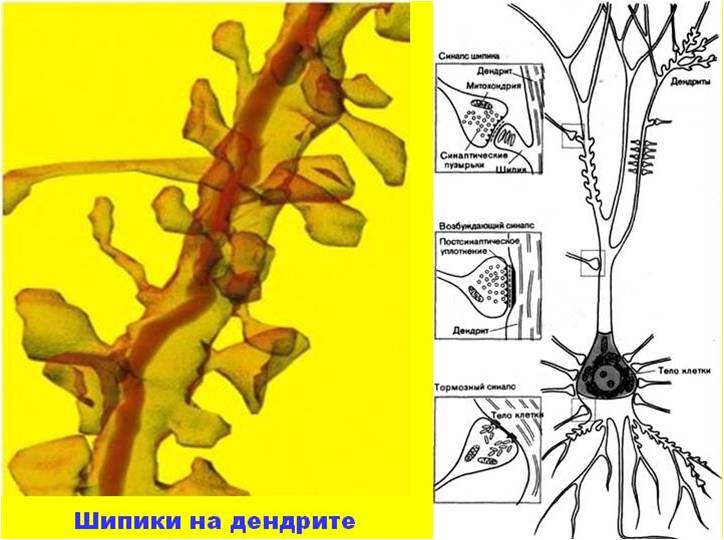 Дендриты — основное воспринимающее поле нейрона. Мембрана дендрита и синаптической части тела клетки способна реагировать на медиаторы, выделяемые аксонными окончаниями, изменением электрического потенциала. Информация дендриту поступает от других нейронов через специализированные контакты-шипики. Чем сложнее функция нервной системы, чем больше разных анализаторов посылают информацию к данной структуре, тем больше «шипиков» на дендритах нейронов. Максимальное количество их содержится на пирамидных нейронах двигательной зоны коры большого мозга и достигает нескольких тысяч. Двигательные пирамидные нейроны получают информацию практически от всех сенсорных систем, ряда подкорковых образований, от ассоциативных систем мозга. Если данный «шипик» или группа «шипиков» длительное время перестает получать информацию, то эти «шипики» исчезают.
Дендриты — основное воспринимающее поле нейрона. Мембрана дендрита и синаптической части тела клетки способна реагировать на медиаторы, выделяемые аксонными окончаниями, изменением электрического потенциала. Информация дендриту поступает от других нейронов через специализированные контакты-шипики. Чем сложнее функция нервной системы, чем больше разных анализаторов посылают информацию к данной структуре, тем больше «шипиков» на дендритах нейронов. Максимальное количество их содержится на пирамидных нейронах двигательной зоны коры большого мозга и достигает нескольких тысяч. Двигательные пирамидные нейроны получают информацию практически от всех сенсорных систем, ряда подкорковых образований, от ассоциативных систем мозга. Если данный «шипик» или группа «шипиков» длительное время перестает получать информацию, то эти «шипики» исчезают.
Аксон представляет собой вырост цитоплазмы, приспособленный для проведения информации, собранной дендритами, переработанной в нейроне и переданной аксону через аксонный холмик — место выхода аксона из нейрона. Аксон данной клетки имеет постоянный диаметр, в большинстве случаев одет в миелиновую оболочку, образованную из глии. Нервные окончания аксонов оказывают трофические влияния на мышцу или клетки других органов. Так, нарушение иннервации мышцы приводит к ее атрофии, усилению распада белков, гибели мышечных волокон. Миелиновые волокна чаще встречаются в двигательных нервах, безмиелиновые преобладают в автономной (вегетативной) нервной системе. В миелинизированных аксонах скорость проведения импульса в разы выше. Считают, что нервное волокно относительно неутомляемо вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения.
Необходимым условием проведения возбуждения в нерве является не просто его анатомическая непрерывность, но и физиологическая целостность. В любом металлическом проводнике электрический ток будет течь до тех пор, пока проводник сохраняет физическую непрерывность. Для нервного «проводника» этого условия недостаточно: нервное волокно должно сохранять также физиологическую целостность. Если нарушить свойства мембраны волокна (перевязка, блокада новокаином, аммиаком и др.), проведение возбуждения по волокну прекращается.
Присутствие в миелиновой оболочке липидов обусловливает их высокое электрическое сопротивление. Обмен липидов в нервной клетке происходит медленно; возбуждение нейрона приводит к уменьшению количества липидов. Обычно после длительной умственной работы, при утомлении количество фосфолипидов в клетке уменьшается. Изначальный недостаток фосфолипидов влияет на процесс миелинизации.
Источником энергии для нервной клетки служит глюкоза крови. Глюкоза расщепляется преимущественно аэробным путем, этим объясняется высокая чувствительность нервных клеток к недостатку кислорода. Даже кратковременное нарушение доставки кислорода кровью может вызвать необратимые изменения в деятельности нервных клеток: в спинном мозгу — через 20 — 30 мин., в стволе головного мозга — через 15 — 20 мин., а в коре больших полушарий — уже через 5 — 6 мин. Увеличение в крови адреналина, активная деятельность организма приводят к увеличению потребления углеводов, следовательно, к повышенному использованию кислороду. Если его поступление недостаточно, возникает гипоксия.
В нервной ткани содержатся соли калия, натрия, кальция, магния и др. роме того, в нейроне имеются различные микроэлементы (например, медь и марганец). Благодаря высокой биологической активности они активируют ферменты. Количество микроэлементов в нейроне зависит от его функционального состояния. Так, при рефлекторном или кофеиновом возбуждении содержание меди, марганца в нейроне резко снижается.
В коре и мозжечке нейроны формируют слои клеток. Каждый слой имеет свою специфическую функцию.
Нейроглия, или глия, — совокупность клеточных элементов нервной ткани, образованная специализированными клетками раз личной формы. Клетки нейроглии заполняют пространства между нейронами, составляя 40% от объема мозга. Глиальные клетки по размеру в 3—4 раза меньше, чем нервные; число их в ЦНС млекопитающих достигает 140 млрд. Клетки глии участвуют в метаболизме, миелинизации нейронов и др. процессах. Показано, что при длительном возбуждении в нейроне высокое содержание белка и нуклеиновых кислот поддерживается за счет клеток глии, в которых их количество соответственно уменьшается.
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния).
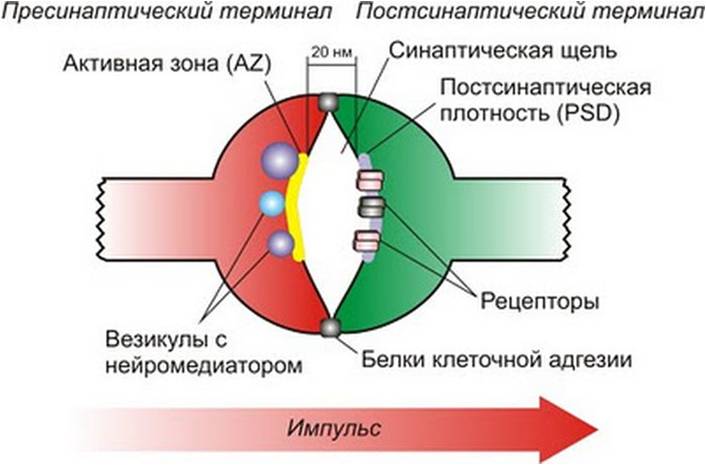 Выполнению функций нейрона способствует синтез веществ-передатчиков — нейромедиаторов (нейротрансмиттеры): ацетилхолина, катехоламинов и др. В синапсах при поступлении сигнала из синаптических пузырьков выделяются медиаторы возбудительные или тормозные. Действуя на постсинаптическую мембрану нейрона, они приводят к изменению ее свойств в области контакта. Суммация этих локальных изменений приводит к изменению внутриклеточного потенциала в сторону его уменьшения (клетка генерирует импульс, передающийся по аксону другим клеткам или органам) или увеличения (нейрон переходит в тормозное состояние). Функционально возбуждение передается только через синапсы. Они выступают как переключатели, которые могут быть еще не включены (до образования долговременной проводимости-памяти) или уже хорошо проводить возбуждение, запуская от пришедшего стимула запомненную связь с ним последующих звеньев, что и обуславливает механизм долговременной памяти. Особенно много их в высших отделах нервной системы и у нейронов с наиболее сложными функциями. При отсутствии активации синапса в течение нескольких дней уже наблюдаются уменьшение размера синаптических пузырьков, запасов медиатора, нарастание длительности синаптической задержки и т. п. Это объясняет постепенное угасание приобретенных навыков (хотя механизм забывания этим не исчерпывается). Понимая суть этого процесса, можно по-новому взглянуть, например, на процесс автоматизации звуков у детей. Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения — сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов — их разрушение, в результате чего полностью нарушается нервная деятельность.
Выполнению функций нейрона способствует синтез веществ-передатчиков — нейромедиаторов (нейротрансмиттеры): ацетилхолина, катехоламинов и др. В синапсах при поступлении сигнала из синаптических пузырьков выделяются медиаторы возбудительные или тормозные. Действуя на постсинаптическую мембрану нейрона, они приводят к изменению ее свойств в области контакта. Суммация этих локальных изменений приводит к изменению внутриклеточного потенциала в сторону его уменьшения (клетка генерирует импульс, передающийся по аксону другим клеткам или органам) или увеличения (нейрон переходит в тормозное состояние). Функционально возбуждение передается только через синапсы. Они выступают как переключатели, которые могут быть еще не включены (до образования долговременной проводимости-памяти) или уже хорошо проводить возбуждение, запуская от пришедшего стимула запомненную связь с ним последующих звеньев, что и обуславливает механизм долговременной памяти. Особенно много их в высших отделах нервной системы и у нейронов с наиболее сложными функциями. При отсутствии активации синапса в течение нескольких дней уже наблюдаются уменьшение размера синаптических пузырьков, запасов медиатора, нарастание длительности синаптической задержки и т. п. Это объясняет постепенное угасание приобретенных навыков (хотя механизм забывания этим не исчерпывается). Понимая суть этого процесса, можно по-новому взглянуть, например, на процесс автоматизации звуков у детей. Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения — сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов — их разрушение, в результате чего полностью нарушается нервная деятельность.
Таким образом, отрицательное влияние на функциональность нейронов оказывают:
1. Отсутствие внешней стимуляции (разрушение митохондрий, меньше синапсов, исчезновение шипиков);
2. Нарушения белкового обмена, отсутствие достаточного количества белков, поступающих с пищей;
3. Гипоксия (отсутствие кислорода препятствует усвоению глюкозы, лишая нейрон энергии);
4. Недостаточность фосфолипидов (быстрее наступает утомляемость, нарушается процесс миелинизации).
Нейроны в коре ГМ в основном представлены двумя видами: звездчатыми и пирамидными. Звездчатые нейроны участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейронов. Пирамидные нейроны осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца — находятся в передней центральной извилине, в так называемой моторной зоне коры. Для коры больших полушарий характерно обилие межнейронных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
Многие нервные клетки обладают способностью к постоянной импульсной активности. Их мембранный потенциал периодически колеблется, то увеличиваясь, то уменьшаясь. Каждый раз, когда его увеличение превышает критический уровень деполяризации (порог возбудимости клетки), возникает импульсный разряд. Такая автоматическая деятельность называется фоновой активностью нейрона. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной активностью. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) - это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи—в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды лишь на новые раздражители, что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.
При ритмических раздражениях вызванная активность нейрона может настроиться на ритм приходящих импульсов (явление усвоения ритма). А.А. Ухтомский доказал важность влияния ритма на работоспособность рефлекторных дуг.
После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозгу (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы и даже дни) и очень велика в коре больших полушарий (до нескольких десятков лет). Различают явные и скрытые следы прежних раздражении. В явной форме сохраняются лишь кратковременные следовые процессы или последствия. Они могут быть одноименными — продолжение возникшего в клетке состояния (тормозное или экзальтационное последействие) или контрастными (возбуждение вслед за торможением, торможение вслед за возбуждением). Непродолжительные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные с биохимическими перестройками в клетках,—в основе долгосрочной памяти.
Кора ГМ имеет электрическую активность. В ЭЭГ различают определенные диапазоны частот, называемые ритмами ЭЭГ (рис. 55). В состоянии относительного покоя чаще всего регистрируется альфа-ритм (8—12 колебаний в 1 сек.), в состоянии активного внимания — бета-ритм (выше 13 колебаний в 1 сек.), при засыпании, некоторых эмоциональных состояниях — тэта-ритм (4—7 колебаний в 1 сек.), при глубоком сне, потере сознания, наркозе — дельта-ритм (1—3 колебания в 1 сек.). В норме амплитуда альфа-ритмов перекрывает остальные колебания. Альфа-ритм исчезает при переходе ко сну.
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе. Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ — быстрой асинхронной активности.
Развитие нервной системы в норме.
Нервная система начинает развиваться, когда большинство женщин еще не подозревает о беременности, а продолжает уже после родов. В постнатальном развитии происходят определенные изменения в плотности расположения нейронов, их объема, ветвления дендритов. С возрастом у человека в мозге число нейронов уменьшается, а число глиальных клеток увеличивается. Развитие мозга определяется двумя факторами: внутренним — генетической программой и внешним — информацией, поступающей извне. Внешняя информация влияет, в том числе, на структурно-морфофункциональную организацию коры.
ЦНС развивается из наружного зародышевого листка, который носит название нервной (мозговой) пластинки. Образование ее относится к 3-й неделе развития эмбриона.
В середине 4-й недели из нее образуется нервная трубка, расширенный конец которой дает начало закладке головного мозга, а вся остальная часть превращается в спинной мозг. Нервная трубка перетяжками делится на 3 мозговых пузыря.
На 5-й неделе из первичных мозговых пузырей образуются 3 главных отдела мозга: передний, средний и ромбовидный. В дальнейшем передний отдел образует конечный и промежуточный мозг, а ромбовидный делится на задний и продолговатый. Из конечного мозга образуются кора и базальные ганглии, а из межуточного мозга — таламус и гипоталамус.
На ранних стадиях развития мозговые полушария имеют гладкую поверхность и тонкую стенку, состоящую из однородных эктодермальных клеток (медуллобластов). После усиленного деления они образуют единый эпендимальный слой. Клетки эпендимной зоны дифференцируются в спонгиобласты и нейробласты — предшественники глиальных клеток и нейронов. В конце 6-й недели нейробласты начинают мигрировать к периферии, образуя плащевой и промежуточный слои, а к концу 2-го месяца (8-я неделя) они передвигаются из плащевого слоя в вышележащий слой, образуя корковую пластинку. В течение 3—4-го месяца корковая пластинка заметно утолщается за счет выселения новых нейробластов, формируя закладку серого вещества. Одновременно идет врастание большого количества отростков нейробластов (нейритов), которые образуют закладку белого вещества.
Разделение первичной коры на отдельные слои начинается на 6—7-м месяце. Процесс этот не заканчивается к концу развития эмбриона и плода, а продолжается и после рождения. Не заканчивается и образование борозд и извилин, которые появляются с 3-го месяца и формируются в постнатальном периоде.
Нейроны созревают первыми и только после завершения нейрогенеза начинается процесс деления и дифференцировки глиальных клеток. На 6—8-й неделе развития нейроны и глиальные клетки, по данным ультраструктурного анализа, имеют все внутриклеточные органеллы, хотя и в меньшем количестве, чем зрелые нейроны взрослого мозга.
Развитие синапса начинается на 4—5-й неделе пренатального периода. Следует подчеркнуть, что синапсы первоначально образуются в избыточном количестве, а затем по мере увеличения специфичности функции отдельных клеток и образуемых ими структур мозга происходит удаление (элиминация) "лишних" контактов. Установление первых межклеточных контактов на 5-й неделе эмбриогенеза служит своего рода границей в онтогенезе нервной клетки — началом ее перехода из стадии нейробласта в стадию юного нейрона. Приводятся данные о том, что уменьшение числа синапсов в коре головного мозга человека происходит и в возрасте между 2 и 16 годами, причем особенно заметные сдвиги такого рода (потеря до 40 % синапсов) имеют место между поздним детским и ранним подростковым периодом. При этом затягивание или ускорение пубертатного периода заканчивается соответственно чрезмерным снижением числа синапсов или, напротив, образованием избытка их (Общая психиатрия/Под ред. А.С. Тиганова).
Если предположить, что полностью развитая нервная система содержит порядка 100 миллиардов (14 в коре) нейронов и что после рождения фактически не добавляется новых нейронов, то можно вычислить, что в период внутриутробного развития нейроны должны образовываться со скоростью более чем 250 000 в минуту. Хотя до сих пор неизвестно, чем включается и выключается механизм размножения в каждой данной области нервной системы, ясно, что сроки, по истечении которых различные популяции клеток перестают делиться, жестко детерминированы. Во многих его областях образуется значительно больше нейронов, чем выживает в последующий период развития. В каждой области, для которой были возможны количественные оценки, обнаружено, что число нейронов регулируется избирательной гибелью клеток, происходящей в предсказуемый период развития (обычно тогда, когда популяция нейронов как целое формирует синаптические связи с тканями-мишенями).
Таким образом, критическими периодами развития ЦНС являются:
3 неделя – образование нервной пластинки.
4 неделя – образование нервной трубки.
5 неделя – образование 3 мозговых отделов из пузырей, образование синапсов.
6-8 неделя – миграция нейробластов.
3-4 месяц - закладка серого вещества, начало образования извилин.
6-7 месяц – начало выделения слоев в коре.
Нейрофизиологические теории, лежащие в основе дефектологии.
 Иррадиация и концентрация нервных процессов. Распространение процесса возбуждения на другие нервные центры называют иррадиацией. Она осуществляется благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг, так что при раздражении одного рецептора возбуждение в принципе может распространяться в центральной нервной системе в любом направлении и на любую нервную клетку. Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Иррадиация через некоторое время сменяется явлением концентрации процессов возбуждения в том же исходном пункте центральной нервной системы. Концентрация происходит в несколько раз медленнее, чем иррадиация нервных процессов. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи — условные рефлексы. Процесс иррадиации тесно связан с торможением, описанным впервые И.М. Сеченовым. Во-первых, процесс торможения ограничивает иррадиацию возбуждения, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов, осуществляя координационную функцию. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
Иррадиация и концентрация нервных процессов. Распространение процесса возбуждения на другие нервные центры называют иррадиацией. Она осуществляется благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг, так что при раздражении одного рецептора возбуждение в принципе может распространяться в центральной нервной системе в любом направлении и на любую нервную клетку. Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Иррадиация через некоторое время сменяется явлением концентрации процессов возбуждения в том же исходном пункте центральной нервной системы. Концентрация происходит в несколько раз медленнее, чем иррадиация нервных процессов. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи — условные рефлексы. Процесс иррадиации тесно связан с торможением, описанным впервые И.М. Сеченовым. Во-первых, процесс торможения ограничивает иррадиацию возбуждения, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов, осуществляя координационную функцию. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
Учение о доминанте. В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров. Термином доминанта был обозначен господствующий очаг возбуждения в центральной нервной системе, определяющий текущую деятельность организма. Основные черты, доминанты следующие: 1) повышенная возбудимость нервных центров, 2) стойкость возбуждения во времени, 3) способность к суммации посторонних раздражении и 4) инерция доминанты. Доминирующий (господствующий) очаг может возникнуть лишь при определенном функциональном состоянии нервных центров. Одним из условий его образования является повышенный уровень возбудимости нервных клеток, который обусловливается различными гуморальными и нервными влияниями (длительными афферентными импульсациями, гормональными перестройками в организме, воздействиями фармакологических веществ, сознательным управлением нервной деятельностью у человека и пр.). Способность стойко поддерживать возбуждение во времени — характерная черта доминанты. Однако далеко не всякий очаг возбуждения становится доминантным. Не сила возбуждения, а способность накапливать и суммировать его превращает нервный центр в доминирующий. Явления суммации лучше всего выражены лишь при умеренном, оптимальном, повышении возбудимости нейронов. Это выражается в том, что доминанта легче всего подкрепляется слабыми раздражениями и гасится сильными. Чем больше нейронов вовлечено в данный очаг возбуждения, тем прочнее доминанта и тем больше она подавляет деятельность других отделов мозга, вызывая в них так называемое сопряженное торможение. Нервные клетки, входящие в доминирующий очаг, расположены не обязательно в одном участке нервной системы. Чаще всего они составляют определенную систему клеток (по А. А. Ухтомскому, «созвездие», или констелляцию, нейронов), расположенных в разных этажах головного и спинного мозга. Объединение большого числа нейронов в одну рабочую систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Следовательно, ритмизация деятельности извне увеличивает шансы возникновения доминанты. Аутогенные тренировки, медитации вызывают устранение доминант, восстанавливая нервную систему. Доминанта является физиологической основой акта внимания. Она определяет характер восприятия раздражении из внешней среды, делая его односторонним, но зато более целеустремленным. При наличии доминанты многие влияния внешней среды остаются вне внимания, но зато более интенсивно улавливаются и анализируются те, которые особенно интересуют человека. Доминанта — мощный фактор отбора биологически и социально наиболее значимых раздражений. Чем выраженнее доминанта, тем больше она тормозит другие текущие рефлексы. Таким образом, из многих степеней свободы выбирается одна.
Три типа функциональных полей мозга. Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову. Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. В коре постцентральной извилины и верхней теменной дольки залегают ядра коркового анализатора чувствительности (температурной, болевой, осязательной, мышечного и сухожильного чувства). Эта зона имеет соматотопическое строение. Ядро двигательного анализатора находится, главным образом, в предцентральной извилине. Размеры проекционных зон различных частей тела зависят не от их действительной величины, а от функционального значения. В коре височной доли находится ядро слухового анализатора. К каждому из полушарий подходят проводящие пути от рецепторов органа слуха как левой, так и правой сторон. Ядро зрительного анализатора располагается на медиальной поверхности затылочной доли. Благодаря близкому расположению ядер обонятельного (лимбическая система, крючок) и вкусового анализаторов (самые нижние отделы коры постцентральной извилины) чувства обоняния и вкуса тесно связаны между собой. Ядра вкусового и обонятельного анализаторов обоих полушарий связаны проводящими путями с рецепторами как левой, так и правой сторон. Вторичные поля служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры — на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария. В третичных полях на основе синтеза всех афферентных раздражении и с Учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
Динамический стереотип. Если раздражители повторяются в определенном порядке, то между ними формируется взаимосвязь, характеризующаяся стереотипной последовательностью возникновения ответных реакций. При этом рефлексы соответствуют не столько данному раздражителю, сколько месту раздражителя в последовательной цепи их. Стереотип внешних проявлений реакций в виде секреции или движения был назван И. П. Павловым динамическим стереотипом. Термин «динамический» подчеркивает функциональный характер этого стереотипа (формирование и закрепление его только после соответствующих упражнений, возможность его переделки, угасание при длительных перерывах, ухудшение при утомлении, сильных эмоциях, заболеваниях и пр.). Переделка стереотипа в некоторых случаях представляет для нервной системы трудный процесс. Чтобы выработать новый стереотип, необходимо сначала угасить старый. Но хорошо закрепленный стереотип трудно подается угашению и может проявляться вновь при возникновении условий, которым он соответствовал.
Теория функциональной системы. Согласно теории П. К. Анохина, именно полезный результат является решающим фактором поведения и для его достижения в нервной системе формируется группа взаимосвязанных нейронов — так называемая функциональная система. Деятельность функциональной системы можно разбить на отдельные последовательные этапы: 1) обработка всех сигналов поступающих из внешней и внутренней среды организма — так называемый афферентный синтез; 2) принятие решения; 3) создание (на основе принятого решения) представления об ожидаемом результате действия и формирование конкретной программы действий для достижения этого результата; 4) анализ полученного результата и уточнение программы действий.
На первом этапе (афферентный синтез) в нервных центрах одновременно взаимодействуют 4 типа поступающих к ним раздражении:
1) пусковая афферентация—сигналы, вызывающие действие;
2) обстановочная афферентация — все остальные внешние раздражения, создающие афферентный фон;
3) мотивация — собственные потребности организма, доминирующие в данный момент: биологические мотивации — жажда, голод, страх и другие, а также высшие мотивации, формирующиеся у человека под влиянием социальных факторов (желание добиться какой-либо цели в жизни, достичь определенного результата в спорте и др.);
4) память—имеющиеся в нервной системе следы прежних раздражении, накопленный опыт.
На основании афферентного синтеза принимается решение («что надо делать») и вырабатывается программа действий («как надо делать»). При этом производится оценка всех возможных путей решения возникших перед организмом задач и ответных реакций организма и выбирается лишь одно необходимое решение. Одновременно в мозгу создается представление о том, какой результат должен быть получен.
Особенности нервной системы – основа компенсации.
Длительное возбуждение нервной клетки может привести к ее гибели. Столь частая гипоксия – снова гибель. Как же мы выживаем?
Пластичность. Есть данные, что одна нервная клетка может заменить девять погибших. Два уровня пластичности: макро- и микроуровень. Макроуровень связан с изменением сетевой структуры мозга, обеспечивающей сообщение между полушариями и между различными областями в пределах каждого полушария. На микроуровне происходят молекулярные изменения в самих нейронах и в синапсах. На том и другом уровне пластичность мозга может проявляться как быстро, так и медленно.
Среди факторов, определяющих восстановительные способности мозга, прежде всего, следует выделить возраст пациента. В отличие от взрослых, у детей после удалений одного из полушарий другое полушарие компенсирует функции удалённого, в том числе и языковые. Второй фактор — длительность воздействия повреждающего агента. Медленно растущая опухоль деформирует ближайшие к ней отделы мозга, но может достигать внушительных размеров, не нарушая функций мозга: в нём успевают включиться компенсаторные механизмы. Однако острое нарушение такого же масштаба чаще всего бывает несовместимо с жизнью. Третий фактор — локализация повреждения мозга. Небольшое по размеру, повреждение может затронуть область плотного скопления нервных волокон, идущих к различным отделам организма, и стать причиной тяжкого недуга. Кровоизлияние в области внутренней капсулы может привести к параличу мышц всей половины тела. Четвёртый фактор — обширность поражения. В целом, чем больше очаг поражения, тем больше выпадений функций мозга. А поскольку основу структурной организации мозга составляет сеть из нейронов, выпадение одного участка сети может затронуть работу других, удалённых участков.
Функциональная специализация разных областей коры мозга определяется их архитектурой. Эта сложившаяся в эволюции специализация служит одним из барьеров для проявления пластичности мозга. Например, при повреждении первичной моторной коры у взрослого человека её функции не могут взять на себя сенсорные области, расположенные с ней по соседству, но прилежащая к ней премоторная зона того же полушария — может. У правшей при нарушении в левом полушарии центра Брока, связанного с речью, активируются не только прилежащие к нему области, но и гомотопическая центру Брока область в правом полушарии. Однако такой сдвиг функций из одного полушария в другое не проходит бесследно: перегрузка участка коры, помогающего повреждённому участку, приводит к ухудшению выполнения его собственных задач. В описанном случае передача речевых функций правому полушарию сопровождается ослаблением у пациента пространственно-зрительного внимания — например, такой человек может частично игнорировать (не воспринимать) левую часть пространства. Идеальным вариантом представляется восстановление функции не за счёт реорганизации поражённого мозга, а за счёт его регенерации. Однако у пластичности мозга есть и отрицательные проявления. Её минус-эффекты определяют многие болезни мозга (например, болезни роста и старения, психические расстройства) (Харченко Е.Н., Клименко М.Н. Пластичность мозга//Химия и жизнь – XXI век).
Спорная способность к нейрогенезу. Клетки нейрогенеза находили у млекопитающих в обонятельных луковицах и коре гиппокампа, которые отвечают главным образом за эмоциональное поведение, реакцию на стресс и регуляцию половых функций млекопитающих. Новые нейроны могут возникать из стволовых клеток даже в мозге взрослого человека (В.Гриневич, Наука и жизнь). В обонятельных луковицах мозга и зубчатой извилине гиппокампа, расположенного на внутренней поверхности височной доли мозга, идёт непрерывное обновление нейронов. Из мозга взрослого человека выделены стволовые клетки, и в лабораторных условиях показано, что они могут дифференцироваться в клетки других органов. Восстановление повреждённого мозга не будет означать полного восстановления прежней личности: гибель нейронов — это невосполнимая утрата прошлого опыта и памяти.
Участие различных участков нервной системы в речевой функции.
Продолговатый мозг. Это продолжение спинного мозга, он расположен на скате затылочной кости. В продолговатом мозгу находится большая группа черепно-мозговых ядер (языкоглоточного, блуждающего, добавочного, подъязычного, нисходящая часть тройничного нерва). На дне IV желудочка в продолговатом мозгу находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха. В продолговатом мозгу находится группа центров моторных рефлексов (жевания, глотания), а также секреторных (слюноотделение). Кроме того, здесь находятся центры некоторых защитных рефлексов: чихания, кашля, мигания, слезоотделения, рвоты. Ретикулярная формация (сетевидная субстанция) - это очень важное образование продолговатого мозга.
Средний мозг. Средний мозг состоит из крыши среднего мозга и ножек мозга. В состав среднего мозга входят четверохолмия, черная субстанция и красные ядра. Срединную его часть занимает ретикулярная формация. Передние бугры четверохолмия представляют собой первичные зрительные центры, а задние бугры—первичные слуховые центры. Ими осуществляют также ряд реакций, являющихся компонентами ориентировочного рефлекса при появлении неожиданных раздражителей (рефлекс «Что такое?»). Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук). Черная субстанция принимает участие во всех движениях организма, регулирует тонус мышц. Нарушение функций и структуры клеток черной субстанции приводит к болезни Паркинсона. Средний мозг выполняет следующие функции: функции движения; сенсорные функции; регулировка продолжительности актов жевания и глотания; обеспечение точных движений рук.
Промежуточный мозг. В состав промежуточного мозга, который является передним концом ствола мозга, входят зрительные бугры — таламус и подбугровая область — гипоталамус.
Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Это так называемые специфические восходящие пути. Они характеризуются соматотопической организацией. Особенно большо

